









Ventricular tachycardia (VT) in the context of structural heart disease is related to patchy or incomplete scar; usually arising from re-entrant circuits which are dependent on surviving channels of activation through scar tissue.1–3 These protected isthmuses are critical for maintaining VT, and an improved understanding of the characteristics of VT isthmuses is important in guiding strategies for VT ablation. VT isthmuses are often unmappable in clinical practice due to non-inducibility, multiple inducible VTs which interrupt the mapping process, and haemodynamic instability during VT.4,5
Historical Data
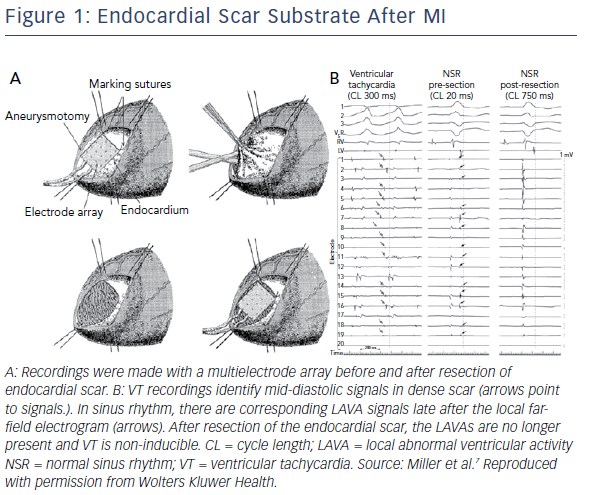
It has long been known that scar-related VT is dependent on poorly-coupled surviving fibres in scarred areas. Early catheter mapping studies identified complex signals with decremental conduction delay which were mid-diastolic during tachycardia.6 Mapping with electrode arrays during surgical ablation of post-infarction VT also identified poorly-coupled signals in areas of dense scar, further demonstrating that these signals were endocardial in origin and separated by dense scar tissue from surviving epicardial myocardium (Figure 1).7
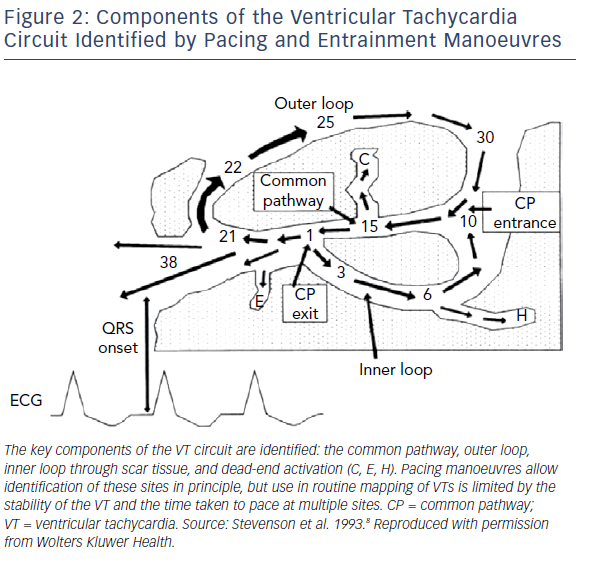
More complex mapping of VT circuits with entrainment manoeuvres to identify concealed entrainment, post-pacing intervals and stimulus-QRS times as well as electrogram (EGM) characteristics, identified the existence of more complex elements of the VT circuit. This included inner and outer loops, a common isthmus and bystander loops or dead ends of activation (Figure 2).8 These techniques are, however, relatively time-consuming because gathering information at each individual point requires entrainment (without terminating the tachycardia) and measurement of intervals. This limits the maximal spatial resolution of the technique and does not allow determination of the anatomical characterisation of the key parts of the VT circuit.
Early electroanatomic mapping systems were used to identify VT circuits with a greater degree of spatial accuracy. This allowed identification of the key characteristics of VT circuits, with identification of typical isthmus orientation and position in post-infarct VTs.1,3 The limited number of recorded points and relatively large sensing bipole used when mapping with an ablation catheter limited the ability of these studies to define the characteristics of VT isthmuses more clearly. Consequently, understanding of the isthmus was limited to site and general orientation, rather than an understanding of more complex details, such as position within the scar, morphology, conduction velocities, and the role of functional and complete block.
High-density Mapping
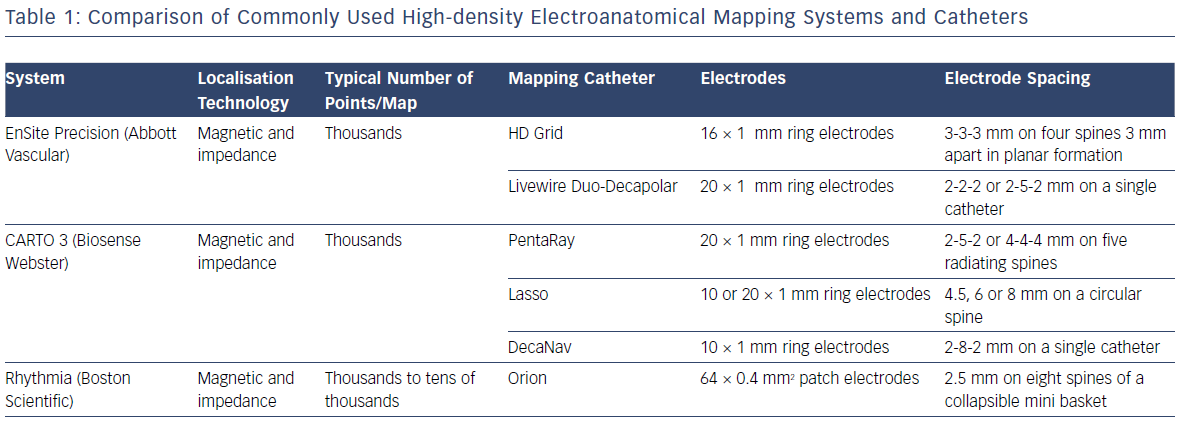
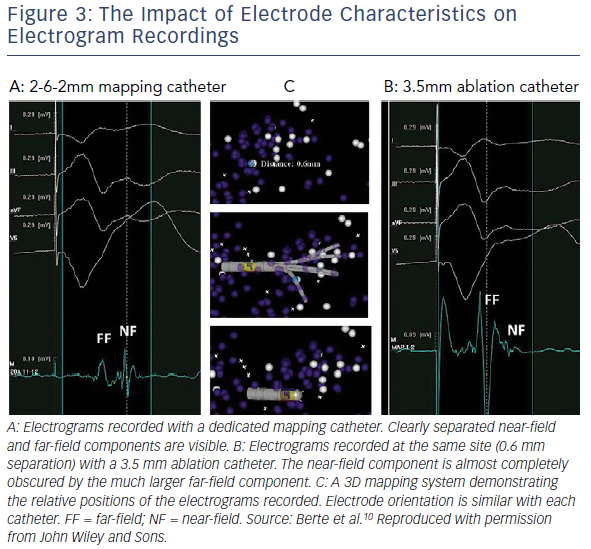
Electroanatomic mapping systems and multipolar mapping systems have been developed which allow collection of greater numbers of EGM from smaller, more closely spaced electrodes (Table 1).9 One of the primary limitations of the techniques described above is that the EGMs were often recorded with a large electrode surface area, particularly in the case of a 3.5 mm ablation catheter tip and relatively large bipolar spacing. Human mapping studies performed with multipolar catheters have clearly demonstrated the effect of increasing bipole distance on the relative sizes of near- and far-field EGMs with clear implications for the ability of human observers and machine algorithms to differentiate the two (Figure 3).10,11 This work has been developed further using the Advisor™ HD Grid mapping catheter (Abbott) in sheep, demonstrating that near-field signals from poorly coupled abnormal fibres are largely unaffected by bipole spacing (as long as the fibre lies between the two poles) but that far-field signals are significantly lessened by reducing the bipole spacing, thereby improving near-field detection.12
A second major limitation of these studies is that the resolution of the mapping techniques is not fine enough to determine the properties of the VT isthmus in detail. A microelectrode study (on a scale of 100 µm) of excised infarcted human papillary muscles identified complex activation patterns in a thin layer of surviving endocardial fibres overlying a densely infarcted zone. Slow conduction in these fibres was the result of activation at near normal speeds on a microscopic level following a zig-zag course through poorly coupled fibres, resulting in overall slowing of conduction.13 Close bipolar spacing is also helpful in this regard, in that the position of the EGM is more accurately determined as signals are recorded from a smaller volume.
The other significant improvement of modern mapping systems lies in the number of EGMs that can be collected, stored and analysed by multipolar recording catheters and improved software. The increase from tens of EGMs in the initial studies to tens of thousands with modern systems has resulted in several major improvements. The resolution of the maps generated has significantly improved. The time taken to generate a map has reduced as multipolar catheters record from areas of myocardium, rather than single points, allowing rapid collection of points over a large area.14,15 By collecting multiple EGMs at each location, outlier signals arising from ectopy, noise or catheter instability can be excluded, improving the quality of the signals recorded.
While these improvements do not allow mapping on the 100 µm scale, to identify activation patterns in individual surviving bundles, they do allow a greater appreciation of VT circuit activation than has been possible before. Several recent studies using small-electrode multipolar mapping catheters and high-density electroanatomic mapping systems have recently been performed in animals and humans, which have improved our understanding of the properties of VT circuits.
Isthmus Architecture
Mapping studies in pigs and humans have confirmed the presence of complex isthmus architecture, as suggested by computer modelling and pacing and entrainment mapping in the era before electroanatomic mapping. A canine left anterior descending (LAD) infarct model mapped with a 192-electrode array identified an isthmus defined by two parallel lines of conduction block.16 This general structure of the common isthmus has been subsequently confirmed in a porcine infarct model, but with multiple entrance and exit sites in 11% of VTs.17 Data from human VT studies confirms this general principle.18 A multicentre study of high-density VT maps found that isthmuses were defined by lines of block, but that multiple entrances and exits were common. Further, in humans, dead ends of activation were also common and regions of activation within dense scar, consistent with inner-loop activation, were also observed.18
This increase in complexity in human studies over animal models is perhaps to be expected. The average time from MI to VT mapping in human studies is far longer, 10–20 years, than the few months possible in an animal model.18,19 The delay in local abnormal ventricular activity (LAVA) and degree of fractionation of EGMs are known to be associated with time from infarction, suggesting a prolonged period of remodelling, which results in increasing architectural complexity with time.20 This complex architecture has important implications in clinical practice.
Both animal and human mapping studies have demonstrated that the same region of poorly-coupled fibres can sustain multiple VTs. Activation passes through the critical VT-supporting area in different directions during different tachycardias, with entrance zones becoming exit zones and vice versa.17,18 Successful ablation in this context requires complete elimination of all potential VT channels. This tendency for several VTs to be possible in an individual patient partly explains why ablation of clinical VTs alone is a less successful strategy than a substrate-based approach, which addresses all potential circuits.21
Conduction Velocity in the Ventricular Tachycardia Circuit
It has long been known that slow conduction, as evidenced by fractionated EGMs, is necessary for VT to be sustained. It also known that conduction velocities in scar tissue are non-uniform and that heterogenous anisotropy results from surviving bundles of myocytes, separated by fibrous tissue and arranged in a mesh-like pattern.22 Previous studies, both surgical and catheter-based, have lacked the resolution to establish whether there is a particular pattern to conduction slowing in the VT circuit.
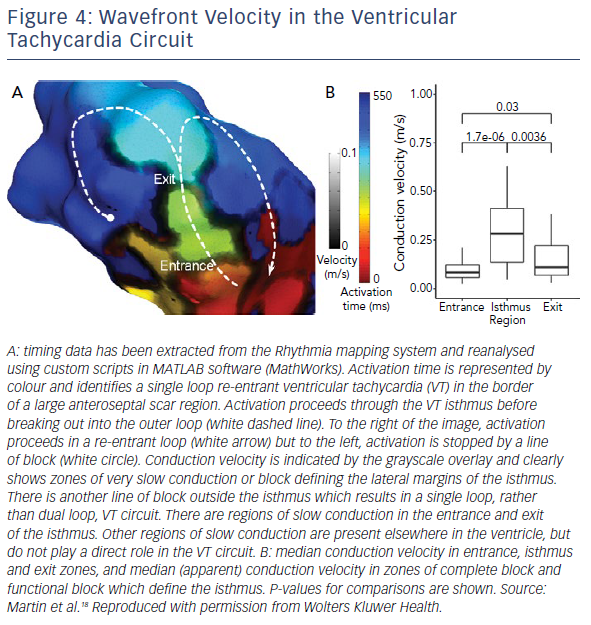
Data from high-density mapping in both animals and humans has identified clear slowing of conduction at VT entrance and exit zones, but relative preservation of conduction velocities in the mid-isthmus (Figure 4).17,18 The absolute values for conduction velocity were higher in a porcine model than in humans, but the velocities recorded in human high-density mapping studies match those seen in surgical mapping studies and the discrepancy likely reflects inter-species difference in myocardial conduction velocity.23
The deceleration of activation wavefronts at entrance and exit zones may be due to one, or more likely a combination, of several factors. Activation in healthy myocardium and in the mid-isthmus is largely linear and orthogonal to fibre orientation. At entrance and exit zones, however, the wavefront curves around the lateral borders of the isthmus, are often perpendicular to fibre orientation. This abrupt change in activation vector as well as increased axial resistivity and myocardial thickness gradient, are likely to contribute to slowing. Further, there is greater slowing of activation in entrance zones due to collision of opposing wavefronts in double-loop circuits.24
The Role of Functional Block and Slow Conduction
As well as slow conduction playing an important role in the entrance and exit zones of the protected part of the VT circuit, regions of slow conduction and/or functional block also seem to be important in defining the VT isthmus. Early animal studies in a canine infarct model identified regions of functional block that were present during tachycardia. High-density mapping in animal and human studies has confirmed this finding. Lines of complete block with well-defined double potentials are observed. However, many VT circuits are also bordered, at least in part, by regions of very slow activation, evidenced by long fractionated EGMs (Figure 5).17,18 Conduction in these lateral borders is sufficiently slow to protect the central isthmus.
Substrate mapping studies in an animal model identified critical sites in zones of maximal wavefront deceleration in sinus rhythm. These sites were located in regions of dense scar (<0.55 mV) and served as anchors of multiple VTs of different configurations and cycle lengths. Multiple VTs are common in clinical practice.25
Implications for Substrate Mapping
The increasing evidence for the role of functional block in VT circuits provides an explanation for the observation of more than one VT breaking out from the same region of scar. A region of slow conduction which acts as a lateral border for one VT may act as the entrance or exit zone for another (Figure 6). The role of functional block results in major limitation of current substrate-mapping approaches, which often rely on a single map generated in either sinus or a paced rhythm.21 Mapping in a rhythm with a cycle length and activation wavefront orientations which differ to the clinical VTs may not identify regions of functional block that are critical to the VT isthmus.26,27 Furthermore, not all LAVA and late potentials lie in critical parts of the VT circuit. Complete substrate elimination, while currently the strategy with the best supporting evidence, may not be entirely necessary for the elimination of all VTs.

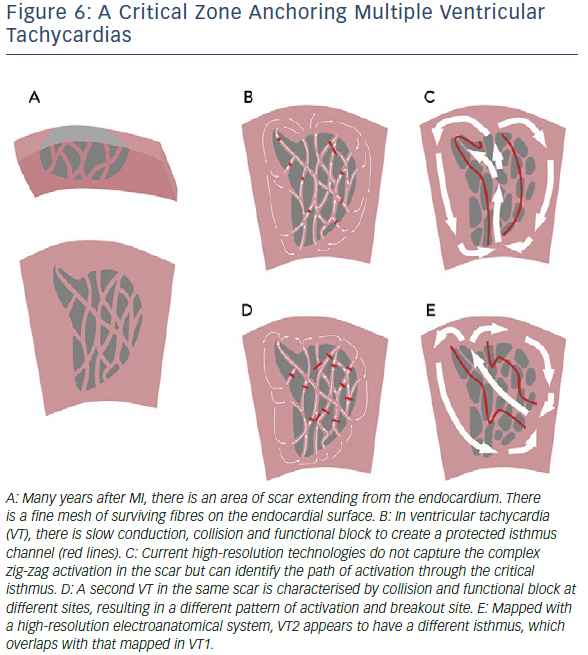
It has been suggested that there may be VT critical zones which serve as the focal point for wavefront slowing and curvature, which can lead to re-entry in multiple different morphologies, leading to multiple VTs.25 Identification of these critical zones in clinical practice may improve the results of substrate mapping by identifying specific areas of LAVA. Strategies to demarcate such zones are needed, but approaches which use pacing from multiple sites and/or different cycle lengths have been shown to improve detection of poorly coupled fibres and are likely to be useful.27 Evidence to support ablation of a targeted subset of LAVA with dynamic poor coupling comes from studies using close-coupled extra-stimuli to identify dynamic increases in EGM duration and latency. Although small, these studies have demonstrated a high rate of non-inducibility and low rate of VT recurrence with a targeted approach.28,29
Implications for Pace Mapping and Entrainment Mapping
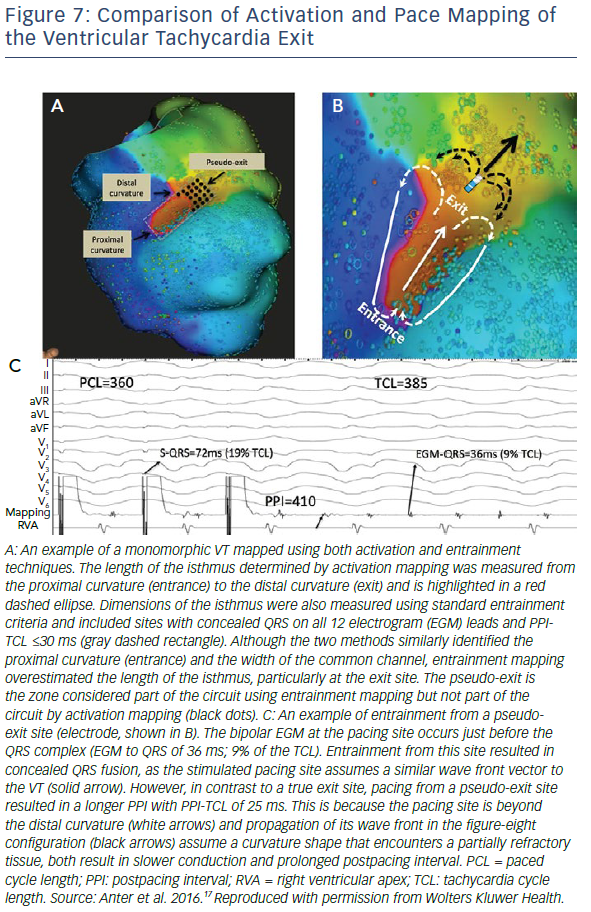
One of the most interesting observations from high resolution studies is the poor correlation of activation mapping and entrainment mapping, particularly in exit zones, at this resolution. Entrainment from sites well beyond the breakout curvature of the activation map still allow concealed entrainment with post-pacing intervals that are consistent with pacing from within the circuit (Figure 7).17 This may be because conduction velocities are relatively slow at the lateral edges of the breakout zone and so activation through this zone contributes very little to the overall morphology of the surface QRS complex. Ablation at these distal sites may fail to terminate tachycardia, however, as activation is still able to break out laterally to healthy myocardium. This phenomenon, although not formally studied, may also have implications for pace mapping. Some studies of pace mapping suggest that the preferred strategy would be to ablate at the entrance site of the identified VT isthmus, which perhaps alleviates this problem.30 Conversely, entrainment and high-density mapping may complement each other. Entrainment remains useful in atrial macroreentry with high-density mapping, and high-density maps may allow a more focused entrainment strategy.31,32
Future Directions
The limitations of current technologies for mapping VT have been reviewed elsewhere, but recent developments in high-resolution mapping are unlikely to be the final improvements in the field.33 The lower limit for electrode size and spacing, beyond which even smaller, more closely-spaced electrodes are no longer helpful, has not been reached. Anatomical studies suggest that the surviving fibres which support tachycardia are about 100 µm in diameter. It is likely that further miniaturisation of recording electrodes to this scale – and perhaps even beyond – will yield more nuanced data on characteristics of VT activation. Any improvement in mapping resolution, however must be matched by localisation accuracy to be clinically useful, as even very large numbers of poorly-located points will not provide an accurate assessment of the isthmus architecture.
The corresponding development of software systems to collect and annotate tens, and possibly hundreds of thousands of EGMs will also continue. Already the number of EGMs collected by current systems defies useful real-time manual analysis in clinical care. Algorithms to display activation throughout the tachycardia cycle length, such as ripple mapping in CARTO (Biosense Webster) or LumiPoint in Rhythmia (Boston Scientific), have been developed and are entering clinical practice.34–37 More sophisticated algorithms to analyse signal characteristics, improving distinction of near-field and far-field signals, and improving annotation of local timing, are in development and will greatly aid the clinician in understanding the VT circuit. Another signal-processing algorithm which shows promise is omnipolar mapping, in which simultaneous analysis of multiple bipolar EGMs allows estimation of wavefront speed and direction from a single beat, assisting in rapid mapping of unstable tachycardias and minimising direction-dependent effects on voltage measurement.
Summary
High-density mapping technologies have improved our understanding of the characteristics of VT isthmuses. These are complex structures with multiple entrances and exits which are defined by a mixture of complete and functional block. These characteristics allow multiple VTs to arise from the same area of substrate. This has clear implications for substrate mapping and ablation strategies, and improved techniques to identify VT critical zones for targeted ablation may improve VT ablation outcomes.38,39
Clinical Perspective
- Ventricular tachycardia (VT) circuits are complex with multiple entrances, exits and dead ends. Tortuous isthmuses are common.
- Local electrogram voltage in the VT isthmus is low, consistent with previous definitions of dense scar.
- Regions of slow conduction play an important role in defining VT isthmuses. These functional elements may make identification of isthmuses in sinus or paced rhythm difficult.
- Several VT circuits may be supported by a single VT critical zone, where a combination of anatomical and functional block supports re-entry.